
Vers le Sommaire
du document
Caractéristiques
biologiques et physico-chimiques du prion ;
Les
hypothèses virologiques ;
Les
hypothèses "protéiques": prions;
molécules
chaperonnes ;
Les
hypothèses "mixtes" : "virinos";
holoprion
Les agents transmissibles non conventionnels sont caractérisés par (28):
- une très importante résistance aux procédés habituels d'inactivation (tableau X ci-dessous)
| Traitement | Conditions | Effet sur la tremblante |
| Procédés physico-chimiques | ||
| NH2OH | 0,1-0,5mM | résistant |
| Psoralen | 10-500µg/ml (50) | résistant |
| Phénol | 100% | inactivation |
| SDS | 1-10% (63) | inactivation |
| ZN2+ | 2 mM (64) | résistant |
| Urée | 3-8M (6) | inactivation |
| Soude | 1 N pendant 1 heure à 20°C (29) | inactivation |
| Hypochlorite de sodium | 2,5% pendant 1 heure à 20°C (6) | inactivation |
| Procédés enzymatiques | ||
| DNase A | 0,1-100µg/ml (52) | résistant |
| DNase | 100µg/ml (52) | résistant |
| Protéinase K | 100µg/ml (62) | inactivation |
| Trypsine | 100µg/ml (62) | inactivation |
| Procédés physiques | ||
| Autoclavage | 136°C pendant 18 min (42) | inactivation |
| Chaleur sèche | 160°C pendant 24 heures (24) | inactivation |
| UV | forte dose | résistant |
- la période d'incubation est longue, excédant souvent 10 ans et pouvant aller jusqu'à 35 ans chez l'espèce humaine;
- les lésions anatomopathologiques sont quasi exclusivement confinées au système nerveux central;
- une absence de signes inflammatoires ou de lésions de démyélinisation dans le système nerveux central;
- l'absence d'anomalies biochimiques ou cellulaires dans le sang ou le liquide céphalo-rachidien;
- l'absence de micro-organismes dans le cerveau des individus infectés lors de l'examen microscopique;
- l'existence d'une modification moléculaire spécifique dans le système nerveux central, à savoir l'accumulation d'une protéine normale de l'hôte, la PrP sous une isoforme pathologique (PrP-res) (65, 64). Les propriétés physico-chimiques de cette protéine sont différentes de celles de l'isoforme normale. L'accumulation de la PrP-res est proportionnelle au titre infectieux;
- la transmissibilité à partir d'ultrafiltrats de broyats cérébraux;
- l'infectivité dépend de la quantité injectée et de la voie prise par
l'infection : la voie intracérébrale est la plus efficace et la voie orale la
moins efficace.
Différentes hypothèses sont actuellement proposées pour expliquer la nature de l'agent responsable de ces encéphalopathies spongiformes transmissibles. On peut les classer en 3 grandes catégories à savoir : les hypothèses virologiques classiques, les hypothèses dites "protéiques" et les hypothèses "mixtes" (d'après D. Dormont, 28).
1) L'hypothèse d'un virus conventionnel inconnu semble a priori peu
probable. En effet ,l'absence de réaction immune, de signes biologiques de
virose, l'absence de particules virales spécifiques et l'insensibilité des
fractions infectieuses aux nucléases (cf. tableau X) ne sont pas en faveur de
cette hypothèse. Les rétrovirus ont été aussi évoqués comme agents étiologiques
potentiels des ESST (46); il est
bien connu que le rétrovirus peut provoquer une spongiose dans le système
nerveux central de la souris. Mais cette hypothèse se heurte aux mêmes problèmes
cités plus haut.
Cependant des travaux récents signalent la présence de
particules pseudovirales dans des cerveaux de hamsters infectés par l'agent de
la tremblante du mouton et dans des cerveaux de patients atteints de MCJ (26). De plus,
l'équipe du professeur Manuelidis indique la présence d'ARN de petite taille (1
000-3 000 bases), potentiellement rétroviral, copurifiant l'infectiosité et
partiellement résistant aux nucléases (47, 48).
2) L'équivalent animal de la viroïde végétale a été avancé au cours des années 70 (25). Mais le manque de sensibilité des ATNC aux nucléases n'est pas compatible avec la sensibilité RNAse des viroïdes.
3) D'autres ont suggéré la présence d'un virus filamenteux.
1) L'hypothèse du prion (64). Cette théorie a été proposée par S.B. Prusiner avant l'identification de la PrP dans les organismes non infectés. De nombreuses données de biologie moléculaire sont en faveur de cette hypothèse :
- les fractions infectieuses purifiées contiennent un seul composant identifiable, la PrP-res, isoforme pathogène de la PrP cellulaire normale. Cette PrP pathogène est partiellement résistante aux protéases;
- la PrP-res s'accumule proportionnellement au titre infectieux sans augmentation des ARN messagers correspondants;
- les souris " Knock-out " dépourvu s de PrP ne sont pas infectables par des fractions de PrP-res homologues;
- l'infectiosité est diminuée par les agents dénaturant les protéines, alors qu'elle n'est pas altérée par des méthodes physico-chimiques dégradant les acides nucléiques;
- les mutations de la PrP peuvent à elles seules entraîner l'apparition d'une encéphalopathie spongiforme subaiguë.
Cet ensemble de données biochimiques et moléculaires a conduit Prusiner et ses collaborateurs à proposer l'hypothèse de "maladie post-transcriptionnelle" (66) qui serait liée à un désordre de l'ingénierie de la synthèse protéique conduisant à l'accumulation d'une protéine normale, sous une forme biochimique pathologique, dans le système nerveux central. Récemment l'analyse de la structure de la PrP et de la PrP-res a permis, par purification spectroscopie infrarouge, d'émettre l'hypothèse d'une conversion des hélices de la PrP en feuillets plissés permettant d'obtenir la PrP-res infectieuse (67), (figure 4).
Cette conversion se déroulerait par formation d'un hétérodimère PrP/PrP-res aboutissant à la formation de deux molécules de PrP-res. Ce processus deviendrait autocatalytique, conduisant à l'accumulation de PrP-res et la mort neuronale.
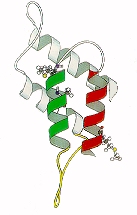

Figure 4. Représentation de la forme normale de la protéine PrP (à gauche) et de la forme infectieuse (à droite), le prion (PrP-res), selon l'équipe de Stantley B. Prusiner (67).
2) La théorie des molécules chaperonnes (39, 45). Ces molécules participeraient au repliement des protéines structurales dans des conformations propres à maintenir leur fonction. On peut supposer que la PrP serait correctement repliée, et que la PrP-res pourrait être une forme au repliement pathologique.
Les hypothèses "mixtes" :
1) A la fin des années 70, les chercheurs britanniques ont proposé le modèle du "virino", particule infectieuse pouvant se répliquer activement ou passivement (41). Ces agents seraient constitués d'une information génétique propre entourée de molécules protéolipidiques de l'hôte, ce qui permettrait d'expliquer leur échappement au système immunitaire. Cette hypothèse expliquerait la variabilité des souches de tremblante, ainsi que la possibilité de mutation.
2) La théorie de l'holoprion, qui prend en compte l'ensemble des données actuelles (77, 78). L'agent infectieux (holoprion) serait l'association d'un petit acide nucléique dont la nature reste à préciser (appelé "coprion") avec la PrP-res ("apoprion"). Dans ce modèle unifié ,le coprion est responsable de la variabilité de la souche et l'apoprion (PrP-res) des phénomènes neuropathologiques qui entraînent la mort des neurones.
D'autres théories, qui impliquent des ARN anti-prion (53), la nucléation (1) ou la maladie toxique (57), ont été proposées. Néanmoins aucune de ces hypothèses ne rend compte de façon définitive et complète des observations cliniques biologiques et épidémiologiques des ESST.
La nature des agents transmissibles non-conventionnels demeure encore aujourd'hui inconnue. Il existe donc des incertitudes majeures, portant sur la nature de l'agent ainsi que sur le niveau précis d'implication de l'origine génétique de l'hôte.
[R] Vers Chapitre précédent; Chapitre suivant; Glossaire; Références bibliographiques; Illustrations
Vers le Sommaire
du document